|
 |
 |
 |
 |
חזרה3 |
(פרק זה מבוסס בעיקר על Di Castri, 1981; מידע שנאסף ממקורות אחרים יצוין בנפרד)
| מבוא |
אקלים ים-תיכוני מופיע בעולם בחמישה אזורים לא רצופים (מפת תפוצת אזורי האקלים הים תיכוני בעולם) - אגן הים התיכון, קליפורניה, צ'ילי, דרום אפריקה ודרום-מערב ודרום אוסטרליה (תנאים ים-תיכוניים מאפיינים חלקים מאזורים אלה אך לא את כל המרחב הגיאוגרפי). אקלים זה מאופיין על ידי קיץ חם, יבש וארוך וחורף לח ומתון. הצומח הטיפוסי לאזורים אלה הם סבכים צפופים יחסית של עצים נמוכים ושיחים, מעוצים, קשי-עלים וירוקי-עד. תצורת צמחייה זו מכונה בארצנו חורש , ובאזורי העולם למיניהם, מכונה בשמות שונים לפי שפת האזור ואופי הצומח. השמות השכיחים ביותר הם: maquis (מקור צרפתי) באגן הים התיכון, macchia באיטליה, chaparral בקליפורניה, matorral בספרד ובצ'ילי, mallee באוסטרליה ו- fynbos או renosterveld בדרום-אפריקה. דווקא באנגלית אין מינוח מיוחד לחורש, ובחוברת זו נעשה שימוש במונח scrubland שפירושו: סבך שיחים קשי-עלים.
לפי שמידע (1980), מוחלפת תצורת הצומח של היער, השליטה באקלימים גשומים (מעל 1,000 מ"מ/שנה), בשתי תצורות צומח באקלימים גשומים למחצה (1,000-300 מ"מ): יער נמוך אך צפוף (חורש, סבך) או יער פארק בו העצים גבוהים אך המרחק ביניהם גדול והנוף הפתוח נשלט, על-פי רוב, על ידי צומח עשבוני.
תנאי האקלים הדומים הביאו להתפתחות צומח דומה באזורים השונים בעולם בהם שולט אקלים ים-תיכוני, במיוחד באגן הים התיכון, קליפורניה וצ'ילי. בכל האזורים הים-תיכוניים הצמחייה השליטה היא עצים ושיחים ירוקי-עד סקלרופיליים (סקלרופילי - גלדני, בעל עלים נוקשים), בתוספת של עשבוניים חד-שנתיים ורב-שנתיים כמרכיבים חשובים. עיקר הצמיחה והפריחה מתרחשים באביב. גם בדרום-מערב אוסטרליה ובדרום אפריקה ישנם אזורים המוגדרים כים-תיכוניים, אך היות והמשקעים בהם מחולקים בצורה שונה, גם פעילות הצמיחה שונה בהשוואה לאזורים הראשונים שהוזכרו (Mooney, Parsons and Kummerow, 1974).
למרות מגבלות המרחב (שטח קטן הנתפס על-ידי צומח ים-תיכוני) והזמן (מערכות אקולוגיות צעירות מבחינה אבולוציונית), יש גיוון רב בהרכב חברות הצמחים ובעלי החיים, הנופים וסוגי הקרקע במערכות אקולוגיות ים-תיכוניות. גיוון זה הוא תוצאה של ההיסטוריה הייחודית של המערכות האקולוגיות המקומיות או האזוריות. חלק מהסיבות שיכולות להסביר גיוון זה מובא להלן:
- מרבית המערכות הים-תיכוניות ממוקמות באזורי מעבר בין מערכות אקולוגיות לחות ויבשות ובין אזורים טרופיים וממוזגים, ולכן רבה בהם ההגירה מהמערכות השכנות.
- המערכות הים-תיכוניות עברו מספר שינויי אקלים חריפים (תקופות הקרח), ובעקבותיהם - חדירות ונסיגות של תצורות צומח שונות לסירוגין. כתוצאה מכך, נמצאים באזור זה צמחים ובעלי חיים ממגוון מקורות ביו-גיאוגרפיים, חלקם שרידים ממערכות אקלימיות אחרות שהסתגלו לתנאים החדשים של יובש קיצי, וחלקם מינים צעירים שהתפתחו מאז הפלייסטוקן כתגובה לאקלים הים-תיכוני.
- מלבד באוסטרליה, רוב המערכות הים-תיכוניות מאופיינות בטופוגרפיה חריפה עם תבליט מודגש, כך שהשטח מחולק, לעיתים קרובות, למגוון בתי גידול רב, העוזר להתפתחות צומח ייחודי.
- כתוצאה מערפל ימי בפסגות ההרים החופיים, השפעות צל-גשם, שיפועים ומפנים שונים, חתירת נחלים גדולים, הימצאות מי-תהום גבוהים והשפעת שריפות חוזרות ונישנות, קיימת נטייה להיווצרות מיני צומח רבים וייחודיים באזורים אלה.
- באזורים הים-תיכוניים השונים קיים מגוון גדול של סוגי קרקע כתוצאה ממספר גורמים: הבדלים גדולים בסלע המקור, הפרשי גובה, שלבים שונים ביצירת קרקע שהתרחשו בהתאם לתנאים פליאו-אקלימיים (פליאו-אקלימי - שייך לאקלימים קדומים) והיסטוריה ארוכה של התיישבות אדם שהשפיעה על תהליכי בליה. ניתן למצוא באזורים הים-תיכוניים סוגים שונים של קרקעות, מצעירות עד עתיקות מאוד, חלקן פוריות מאוד וחלקן עניות ביותר בחומרי מזון, בעיקר זרחן וחנקן.
- רוב המערכות האקולוגיות באזור הים-תיכוני נמצאות תחת ההשפעה המתמשכת של פעילות האדם, ולכן מראות מאפיינים שונים של מבנה ותיפקוד (וכתוצאה מכך הן מצויות בשלבים סוקצסיוניים [סוקצסיה - תהליך אקולוגי, על-פי רוב ממושך, שבמהלכו משתנה הצומח מחברות חלוץ, הראשונות להתיישבות בשטח אחרי הפרעה אך חסרות התאמות מפותחות לחיים בו, עד לצומח השיא (קלימקס), המותאם בצורה הטובה ביותר לתנאים המקומיים] שונים). חלק מהמערכות הן תוצר של דגרדציה (התנוונות) של מערכות מורכבות יותר; אחרות נמצאות בשלב התאוששות לאחר שהשפעת האדם נפסקה, ואחרות יכולות להיות במצב המתקרב לתנאי שיא ההתפתחות - קלימקס.
כתוצאה מהמגוון במרחב ובזמן, ניתן למצוא באזורים הים-תיכוניים פסיפס של תת-מערכות אקולוגיות שונות בשטח קטן. מינים ממשפחות וקבוצות שונות (בעלי מקור פילוגנטי [מבטא את הפילוגניזה – ההיסטוריה ההתפתחותית-אבולוציונית של קבוצת אורגניזמים] שונה) וממקורות גיאוגרפיים שונים, ומינים בעלי אסטרטגיה אבולוציונית (התאמות לתנאי סביבה) ועמידות אקולוגית שונה, יכולים להופיע באותה חברה; שימושי קרקע שונים יכולים להתקיים ולהצליח באותו שטח; טקסונים (טקסון - ביטוי כולל ליחידת מיון בסיסטמטיקה [מזן ועד מערכת]) שונים - בעיקר של צמחים עשבוניים ובעלי חיים בקרקע – מגיעים לשיא המגוון הגנטי באזורים אלה, למרות התנאים הקשים של האקלים הים-תיכוני.
למרות השוני הקיים בתוך כל אזור והעובדה שבכל אחד מהאזורים הים-תיכוניים של העולם התפתחו צומח וחי ייחודיים, מבודדים, וממקורות פילוגנטיים שונים, ישנו גם דמיון רב בין האזורים הללו. דמיון זה מתבטא באקלים, בתצורת הצומח הטבעית השליטה, באופי הנוף, בהתאמות הצומח והחי לתנאים הסביבתיים ולגורמים המגבילים, ובבעיות הממשק וניהול המשאבים הביולוגים של האקוסיסטמה הים-תיכונית באזורים הים-תיכוניים בעולם.
לא בכדי נעשו מאמצים מדעיים לא מבוטלים לאפיין את הדמיון שבין הצומח הים-תיכוני (ראה סעיף 5 בפרק זה) באזורים השונים (Mooney and Dunn, 1970; Cody and Mooney, 1978; Tenhunen et al., 1987; Specht, 1969; Shmida, 1981; Naveh, 1967; Di Castri, Goodall and Specht, 1981; Di Castri and Mooney, 1973;) אבל גם ריבוי עבודות אלה השאיר את חידת הקונברגנציה (התפתחות אבולוציונית המתרחשת במסלולים התפתחותיים נפרדים ושונים והמגיעה לתוצאה אקולוגית, מורפולוגית או התנהגותית דומה) של הצומח הים-תיכוני בעולם בלתי פתורה (ראה סעיף 6 בפרק זה).
| החורש הים-תיכוני - הגדרה |
מבין תצורות הצומח הנפוצות באזורים הים-תיכוניים, אנו מכירים את הבתה המורכבת בעיקר מבני שיח וצומח עשבוני, את הגריגה הנשלטת על-ידי שיחים בגודל בינוני (עד 2.0 מ'), את יער הפארק המורכב עצים מפותחים (עם גזע יחיד) הגדלים בתפזורת, וביניהם צומח עשבוני, ואת החורש המורכב שיחים גבוהים ועצים נמוכים (על-פי רוב, עם מספר גזעים) בדרגות כיסוי וגובה שונות. תנאי בית הגידול והשפעת פעולות האדם יצרו ואריאציות שונות של תצורות צומח אלה. דפני (1977) הגדיר יער כתצורת צומח של עצים בעלי גזע מרכזי שגובהם מעל 3 מטר, ואילו חורש הוגדר כתצורת צומח של עצים בגובה 5-3 מ', בעלי מספר גזעים.
חוקרים רבים עסקו בתיאור החורש הים-תיכוני, אך עדיין קשה מאוד לתת הגדרה אחת המכסה את כל התצורות הצמחיות השונות הנכללות במושג "חורשים ים-תיכוניים". ננסה לאחד ולסכם את ההגדרות והתיאורים של כמה מהם, ולנסות ולקבל הגדרה אחת שאליה נתכוון בהמשך, בהזכירנו את המושג "חורש ים-תיכוני".
Specht מצטט (1969) את Schimper מ-1903 שטען שלמרות ההבדלים במינים, בסוגים ובמשפחות הצמחים, יש עדיין דמיון רב בצורה ובמורפולגיה של הצומח באזורי אקלים ים-תיכוניים בכל העולם. לפי Schimper, תצורת הצומח האופיינית לאזורים אלה מורכבת מצמחים מעוצים, רבי-גזעים, ירוקי-עד, בעלי עלים נוקשים. באותם מקומות שבהם הצומח הושפע רק מעט על ידי האדם, מופיעה חברת החורש: עצים נמוכים, קשי-עלים, בעלי מספר גזעים קצרים וענפים מפותלים, לעיתים קרובות מתפרסים. בשכבה התחתונה נוכחים עצים קטנים ננסיים או שיחים גבוהים (פאנרופיטים) (פאנרופיט – שיחים גבוהים ועצים בעלי ניצני התחדשות בגובה מעל 80 ס"מ מעל פני הקרקע), בני-שיח (כאמפיטים) (כאמפיט – שיחים נמוכים ובני שיח בעלי ניצני התחדשות קרובים לקרקע, ובכל מקרה לא יותר מ-80 ס"מ מעל פני הקרקע) וצמחי בצל ופקעת (גיאופיטים) (גיאופיט – צמח רב-שנתי בעל ניצן התחדשות המצוי מתחת לפני הקרקע. בעל פקעת בצל, פקעת-שורש או קנה-שורש); תיתכן גם נוכחות של מטפסים דקים מעוצים; מינים חד-שנתיים נפוצים באגן הים התיכון, אך נדירים יותר בשאר האזורים הים-תיכוניים. שימפר כינה צומח זה “sclerophyllous woodland” ותיאר דוגמאות מחמשת אזורי האקלים הים-תיכוני.
בשלב מאוחר יותר, קבע ( Warming (1909 ארבע תצורות צומח במסגרת הגדרותיו של Schimper, המקובלות גם על חוקרים רבים נוספים:
- ערבה שיחית (dwarf-shrub steppe) - תצורת צומח נמוכה המורכבת בעיקר מבני שיח וצומח עשבוני. חלק מהשיחים הנמוכים בתצורה זו הם נשירי קיץ או מחליפי עלי חורף בעלי קיץ. תצורה זו זכתה לשם התנ"כי בתה (batha), שהוענק לה על-ידי איג (Eig, 1927).
- גריגה (garrigue) - תצורה פתוחה של שיחים קטנים, ירוקי-עד בעלי עלים גלדניים, בגובה 1.5-0.5 מ' עם קומה תחתונה המורכבת מבני שיח סקלרופיליים (סקלרופילי – גלדני, בעל עלים נוקשים), עשבים רב-שנתיים וגיאופיטים. תצורה זו נפוצה סביב הים התיכון ומקורה, כנראה, בהרס חורשים או יערות. לפי הטרמינולוגיה הצרפתית, הגריגה היא תצורת צומח ים-תיכונית מעוצה שמאפייניה דומים לאלה שתוארו לעיל, אך תפוצתה מוגבלת לקרקעות המתפתחות על סלעים גירניים.
- טומיירס (tomillares) - תצורה מצויה בספרד, דומה לגריגה, אך נשלטת על ידי בת-קורנית (Thymus).
- חורש (maquis) - תצורה צפופה של עצים קטנים ושיחים ירוקי-עד, בעלי עלים גלדניים שגובהם 3-1 מ' עם שכבה תחתונה של שיחים נמוכים קשי-עלים בגובה 1.5-0.5 מ', עשבים רב-שנתיים וגיאופיטים. תצורה זו נפוצה בסביבות הים התיכון, ויש לה מקבילות בדרום-אפריקה במושבת הכף (שם מקומי – fynbos), בצ'ילי (espinal), בקליפורניה (chaparral) ובדרום אוסטרליה (mallee). בצרפת, לעומת זאת, מוגדר המאקי (maqui), כחורש המתפתח רק על קרקעות חמוצות (שמקורן בסלעי יסוד או מטמורפיים). (השמות המובאים כאן הופיעו כבר אצל Warming, וחוקרים מאוחרים השתמשו בחלק מהמקרים בשמות שונים.)
למעשה, הגדרה זו של החורש מקובלת על כל החוקרים העוסקים בצמחייה הים-תיכונית, עם שינויים קלים הקשורים, בדרך כלל, לאזור הספציפי הנחקר (ראה למשל, Shmida and Barbour, 1982; Zohary, 1982; Di Castri, 1981; שמידע 1980, א ו-ב; ויזל, פולק וכהן, 1975; ויזל, 1984).
הגורמים האקולוגיים העיקריים הקובעים את התפתחות הצומח הים-תיכוני הם: יובש קיצי וזמינות חומרי מזון בקרקע. בהתאם, לאורך גרדיאנטים של יובש עולה (או בבתי גידול מופרעים יותר) הצמחייה נעשית נמוכה יותר (כ-1 מ' גובה) ופתוחה או מפוזרת יותר. לפעמים (במיוחד בצ'ילי וקליפורניה) יש יחס גבוה של שיחים נשירי קיץ או ננסיים - זאת התצורה המכונה בארץ בתה, jaral בצי'לי, phrygana ביוון, nenosterbos בדרום-אפריקה ו- costal sage בקליפורניה. לאורך גרדיאנטים של לחות עולה, יופיע חורש מפותח ואח"כ יער פארק של אורן (קליפורניה ומרכז הים התיכון).
שמידע (Shmida, 1981), מציע סכמה המקשרת בין תצורות הצומח ומפל המשקעים: באזור המקבל מעל 1,000 מ"מ גשם, ישלוט יער נשיר מעורב, ב-1,000 מ"מ ופחות - יער ירוק-עד מעורב. במשטר של 1,000-800 מ"מ, ניתן למצוא יער פארק של אלון-אורן או matorral גבוה וסבוך. באזור בו יורדים 600-500 מ"מ גשם בממוצע, יתפתח חורש או יער פארק נמוך. משטר יבש יותר יגרום להשתלטות תצורת צומח נמוכה ופתוחה יותר, שיחית באופיה.
בבתי גידול ממוזגים יותר, החורש הים-תיכוני יכול להפוך ליער של עצים רחבי עלים, ירוקי-עד, סקלרופיליים שבו נעלמת הקומה התחתונה. כאשר האדמה ענייה במיוחד בחומרי מזון, בעיקר על קרקעות חול שטופות מאוד או קרקעות חומציות, מתפתחת במקום החורש תצורת צומח מעוצה אחרת: בתת אברשים – heathlands (אוסטרליה ודרום אפריקה). ככל שפוריות הקרקע עולה, מהווים צמחים עשבוניים מרכיב חשוב יותר בחורש הים-תיכוני, ואז ניכרת נטייה לכיוון יצירת נוף סאוואנה, יער פארק או יער פתוח.
גורם נוסף בבקרה האקולוגית באקלים הים-תיכוני, שאינו זוכה לעיתים קרובות להתייחסות נאותה, הוא טמפרטורות נמוכות בחורף (ראה פרק אקלים – איור 1). הגורם התרמלי הוא כנראה בעל משמעות רבה יותר באגן הים התיכון מאשר באזורים ים-תיכוניים אחרים, היות ובאזור זה, ובעיקר באזורים רחוקים מחוף הים או בבתי גידול רמים טופוגרפית, מתפתחים תנאי אקלים הדומים לתנאים יבשתיים המאופיינים במעברי טמפרטורה חדים בין היום ללילה ובין הקיץ לחורף. עם זאת, גם באזורים ים-תיכוניים אחרים יש חורשים הרריים באזורי המעבר ליערות מחטניים ו/או יערות נשירים.
האדם מילא תפקיד מפתח ביצירה ובהתפתחות של כמה מיחידות הצומח הים-תיכוני. זאת על ידי התערבות מסיבית בנוף, בפעולות כמו: רעייה חזקה, כריתה, שריפה ועיבוד חקלאי שגרמו להיעלמות מיני צמחים וסחיפת קרקע. עקב כך, הוחלפו התצורות הקודמות בתצורות שיחיות מפוזרות וקוצניות. בעיקרון ניתן לומר, שככל שהתערבות האדם חזקה יותר תצורת הצומח המתקבלת נמוכה ופתוחה יותר. עם הסרת גורמי ההפרעה הולך הצומח ומתאושש, דבר הבא לידי ביטוי בהגדלת כיסוי הצומח המעוצה ובעליית הגובה של תצורת הצומח.
ההפרדה בין החורש לבין תצורות צומח אחרות, המופיעות גם הן באקלים ים-תיכוני, כגון יערות קשי-עלים, יערות מחטניים ובתות, היא מלאכותית במידת מה. החורש הוא, במקרים רבים, שלב סוקצסיוני במגמת התאוששות או נסיגה, הקשור במעבר רציף למערכות אקולוגיות אחרות, ומהווה כעין שלב ביניים שקשה לקבוע את משמעותו. רוב בעלי החיים למשל, אינם מבחינים בתפוצתם ופעילותם בגבול שבין החורש ליחידות אקולוגיות אחרות.
פעילות האדם קיימת בכל הקשת של צורות הצומח באזור זה. למעשה, אין מערכת אקולוגית יחידה שיכולה להחשב כייחודית לסביבה הים-תיכונית המאופיינת על-ידי מגוון נרחב של תנאים סביבתיים ביוטיים וא-ביוטיים.
| תפוצת החורשים הים-תיכוניים |
תפרוסת האזורים הים-תיכוניים בעולם לפי ( Di Castri (1981, היא למעשה גם מפת תפוצת החורשים הים-תיכוניים בעולם. בקריאת המפה חייבות להיות מובאות בחשבון מספר הסתייגויות:
- מפה בקנה מידה כזה היא בהכרח כוללנית. אזורים מסוימים שבהם החורש הים-תיכוני הוא צר מאוד, כגון לאורך חלק מהחופים, מוגזמים, במידה מסוימת, בתפרוסתם במפה.
- המפה אינה משקפת את העובדה שרוב החורשים הים-תיכוניים הם בעלי תפוצה בלתי רצופה, גם בתוך אזורים מצומצמים, כתוצאה משינויים טופוגרפיים, מיקרו-אקלימיים או של קרקע. כמו כן, אזורים נרחבים הושפעו רבות על ידי האדם - למשל, מרבית שטחי אגן הים התיכון.
- במפה מופיעים לא רק חורשים ירוקי-עד סקלרופיליים, אלא גם תצורות צומח אחרות, פתוחות ונמוכות יותר, בעלות צמחים נשירי קיץ וסוקולנטים (סוקולנטי – בשרני,עסיסי), שטחים עשבוניים וסאוואנות וגם יערות פתוחים. כמו כן, בדרום אפריקה ואוסטרליה אין אפשרות של הבחנה ברורה בין חורשים ותצורות צומח אחרות הנפוצות באזור הים-תיכוני.
בנוסף לכך, המפה אינה מתיימרת להראות את השטחים המדויקים בהם חורשים ים-תיכוניים אכן נמצאים למעשה, אלא מראה בעיקר שטחים שבהם יש פוטנציאל להתפתחות חורשים כאלה, גם אם תפוצתם כיום מאוד לא רציפה. בכל זאת, ניתן לעשות מספר הכללות על סמך המפה:
- חורשים ים-תיכוניים קיימים, במידה מסוימת, בכל חמשת היבשות, בעיקר בין קווי הרוחב ˚30 ו- ˚40, מצפון ומדרום לקו המשווה.
- רוב התצורות הים-תיכוניות קיימות בחופים המערביים של היבשות. יש חדירה מזרחה רק באגן הים התיכון ובאוסטרליה. בקליפורניה וצ'ילי, הגבול המזרחי מיוצג על ידי שרשרות הרים מפותחות, ובדרום אפריקה ואוסטרליה על ידי הגשם הקיצי של החופים המזרחיים.
- החורשים הים-תיכוניים בקליפורניה, צ'ילי ואגן הים התיכון נמצאים בערך במרכז של מסות יבשתיות גדולות. למשל, אגן הים התיכון ממוקם במרכז של מסה יבשתית כמעט רצופה המיוצגת על ידי אירופה, אסיה ואפריקה.
כ-60% משטחי החורשים הים-תיכוניים בעולם מופיעים באגן הים התיכון. הגורם לכך הוא נוכחות הים התיכון המאפשרת את החדירה המזרחית ביותר של האקלים הים-תיכוני. בנוסף, קיים שטח גדול של חוף (סביב האוקיינוס האטלנטי, הים התיכון והים השחור) ומספר גדול של איים בים התיכון (בסך הכל, יותר מ-40,000 ק"מ של חופים) המגדילים את האפשרויות למצוא תנאים נוחים להתפתחות חורשים גם באזורים שמחוץ לקווי הרוחב האופייניים להם. עם זאת, מסתבר שמבחינת שטח רצוף של חורשים גלדניים ירוקי-עד, קליפורניה נמצאת במקום הראשון.
| חקר מערכות אקולוגיות ים-תיכוניות |
מחקרים רבים, הכוללים גישות רבות ושונות, נעשו באזור הים-תיכוני ברמת המערכות האקולוגיות הכלולות בו. לפי , 1981Dבשנת i Castri ו- Quezel בשנת 1981a ניתן להבחין בשלוש גישות עיקריות לנושא:
- הגישה הפיטו-אקולוגית, ששורשיה נעוצים בעבודתם של Gaussen, Braun.-Blanquet ובעיקר Emberger, בחצי הראשון של המאה העשרים. שני העקרונות של גישה זו הם איפיון מפורט של הביו-אקלימים הים-תיכוניים השונים ותיאור מדויק של חברות הצמחים. השיטה מתבססת בעיקר על ניתוח הצומח, ואינה כוללת תיאור אקולוגי של הקרקע או החי. היא מופעלת על יחידות שטח גדולות יחסית ומקילה על אפשרויות היישום וההתחשבות בהשפעת הגורם האנושי. האסכולה הפיטו-אקולוגית או הפיטו-סוציולוגית נמצאת בשימוש נרחב, בעיקר בכל אזור אגן הים התיכון.
- הגישה הפונקציונלית הקשורה לאקולוגיה של מאזן אנרגיה (אקולוגיה של יצרנות). זאת גישה אינטגרטיבית הכוללת מחד גיסא, ניטור של ייצור ראשוני, ביומסה, ייצור שניוני, פירוק (decomposition) ומסלולי זרימת אנרגיה ומיחזור חומרי מזון, ומאידך גיסא, מדידת משתנים סביבתיים בלתי תלויים, כגון: טמפרטורה, משקעים וקרינה. בסופו של דבר, אמורים מודלים של המערכת האקולוגית לספק כלים לחיזוי ההשפעות של אלטרנטיבות ממשק שונות, שמטרתן להגיע לייצור ביולוגי מקסימלי עם דגרדציה מינימלית של הסביבה. המחקרים נערכים על שטחים מצומצמים יחסית, שבהם מתרכז מאמץ אינטנסיבי מאוד. אחת ממגבלות השיטה היא המחסור בראיות אמפיריות על אפשרות השלכת המסקנות מאתר אחד לאתר אחר, או על הכללת המסקנות לשימוש ברמה אזורית או בין-יבשתית. בעיה זו גדולה במיוחד במערכות אקולוגיות ים-תיכוניות, כתוצאה מהשוני הרב הקיים בתוך המערכות ומבנה הפסיפס של הצומח והקרקע. יש גם פערים במידע שנאסף לגבי האזורים הים-תיכוניים בעולם, למשל: אין כמעט מידע על מטבוליזם של אנרגיה וחומרי מזון בקרקע או הפעילות של חסרי חוליות באזורים אלה.
דוגמה מקיפה למחקר בגישה זו הוא ניתוח המבנה והדינמיקה של הפריגנה (בתה) היוונית, שכלל לימוד כמותי של הביומסה הצמחית לאורך עונת הצימוח, אוכלוסיות המיקרואורגניזמים בקרקע, תכולת חנקן וכלורופיל בצמחים וחלקיהם, ייצור ראשוני של האקוסיסטמה, תחלופת השורשים, אינדקס שטח העלים, משקל העלים ושטח הפחמן והחנקן במערכת (Margaris, 1976). - הגישה האבולוציונית. גישה זו מנסה להגדיר עבור חלקי מפתח של המערכת האקולוגית את: (א) האסטרטגיות האבולוציוניות (התאמות לתנאי הסביבה) העיקריות ו-(ב) הכוחות הסלקטיביים הגורמים להתפתחות האסטרטגיות הנ"ל. היא גם מנסה לבדוק את תוצאות המחקר ב: (א) אזורים סמוכים לאורך גרדיאנטים סביבתיים של יובש, לחות או קור - כלומר שינויים אקלימיים הפועלים על מינים קרובים ו-(ב) אזורים מרוחקים בעלי תנאים א-ביוטיים דומים עם מינים רחוקים ממשפחות/קבוצות שונות. גישה זו מביאה בחשבון גורמים היסטוריים ושינויים גיאוגרפיים. הסקלה של יחידת המחקר קטנה מזו שבגישה הפיטו-אקולוגית, אך חוזרת בווריאנטים שונים של אקלים וגיאוגרפיה. לסיכום, הגישה האבולוציונית פחות יקרה ומתוחכמת מהגישה של מאזן אנרגטי, אך עוד לא נבדקו בה היישומים האפשריים לממשק. גישה זו התפתחה בעיקר בהשוואות של אבולוציה קונברגנטית (קונברגנציה – התפתחות אבולוציונית המתרחשת במסלולים התפתחותיים נפרדים ושונים והמגיעה לתוצאה אקולוגית, מורפולוגית או התנהגותית דומה) במערכות האקולוגיות הים-תיכוניות בצ'ילי וקליפורניה (Di Castri and Mooney, 1973), תוך ניסיון להגדיר אדפטציות דומות לתנאים דומים.
הבעיות העיקריות הקיימות בשימוש בגישות השונות הוא היעדר דו-שיח והיזון חוזר ביניהן. אין מספיק חילופי נתונים בין מחקרים שנערכו בשיטות השונות כדי לעשות בהם שימוש בבניית תמונה כוללת. בעיות אלה נובעות ממספר גורמים:
- קשיי שפה - סקר ספרותי מראה שמאמרים רבים שנכתבו בצרפתית, ספרדית ואיטלקית אינם נכללים בקבצי מידע בינלאומיים.
- בעיות ניסוח - חוקרים שונים משתמשים במונחים רבים ושונים המקשים מאוד על השוואה בין ממצאיהם.
- בעיות מתודולוגיות ומושגיות - בעיקר בהקשר לקנה המידה של שטחי המחקר (כפי שפורט לעיל) ובפירושים שונים הניתנים למושג חברת הקלימקס במערכות אקולוגיות ים-תיכוניות.
- פערים בידע - בתחומים מסוימים (למשל נושא הקרקע) חסר מידע בסיסי. בתחומים אחרים, המידע מאזורים ים-תיכוניים שונים אינו אחיד ולכן קשה לערוך השוואות.
- בעיות כלליות הקשורות לעצם מהותה של המערכת האקולוגית הים-תיכונית המורכבת, והשונות הרבה שבתוכה.
| דמיון ושוני בין תצורות חורש בעולם |
האזורים השונים של האקלים הים-תיכוני בעולם מראים דמיון ברור בתכונות היסודיות, אך לכל אחד ישנם גם מאפיינים ייחודיים. מפה 3 מציגה את הדמיון והשוני במאפיינים האקלימיים היסודיים של חמשת האזורים הים-תיכוניים. כללית ניתן לומר שהצומח במוקדי ההתפתחות השונים של חורש ים-תיכוני בעולם מורכב מכיסוי צפוף של צמחים רב-שנתיים ירוקי-עד שגובהם הממוצע 3-2 מ', וביניהם קרחות או כתמים חשופים בהיקף מוגבל.
איור 3 מציג השוואה פיזיוגנומית (של המבנה הפיזי) בין תצורות החורש מאזורים שונים בעולם (Cody and Mooney, 1978).

טבלה 1 מציגה השוואה של ההרכב הפלוריסטי ותצורות החיים ב- 4 מוקדי חורש בעולם (Cody and Mooney – ממקורות שונים). לפי השוואה זו, בדרום אפריקה קיים הצומח הים-תיכוני העשיר והמגוון ביותר מבחינה פלוריסטית.

גם בצ'ילי צומח ים-תיכוני מגוון, בעוד שבקליפורניה וסרדיניה נמצאו התצורות העניות יותר מבחינת מגוון המינים. בסך הכל, הדמיון בהרכב הפלוריסטי מוגבל. שכיחות הצומח המעוצה דומה בסרדיניה, דרום-אפריקה וקליפורניה (50%-35% מסה"כ הצומח) אך נמוכה יותר (20%) בצ'ילי. דמיון נמצא גם במאפיינים אקולוגיים של חברות החורש באתרים השונים (טבלה 2).

| קונברגנציה בין תצורות חורש בעולם |
המושג קונברגנציה (convergence) או התפתחות קונברגנטית (גם התפתחות אנלוגית) בא לתאר התפתחות תכונות פנוטיפיות (פנוטיפי – תכונות שאינן נקבעות גנטית, אלא מתפתחות כתגובה לתנאים מיידיים) דומות באורגניזמים שאין ביניהם קשר סיסטמתי או פילוגנטי (פילוגנטי - מבטא את הפילוגניזה – ההיסטוריה ההתפתחותית-אבולוציונית של קבוצת אורגניזמים) (אינם קרובים מבחינת מעמדם הביולוגי) ו/או קשר גיאוגרפי. זאת בניגוד להומולוגיה, שהיא התפתחות בקווים דומים, אשר להם מקור (אב קדמון) משותף (קרויה גם התפתחות במקביל – parallel evolution). מקובל להניח שהתפתחות קונברגנטית מתרחשת כאשר אורגניזמים שונים עומדים, באתרים גיאוגרפיים נפרדים, בפני תנאים אקולוגיים דומים. במצב כזה, עשויות להתפתח התאמות דומות אצל כל האורגניזם בנפרד וללא תלות הדדית. בצמחים, הקונברגנציה אמורה להתבטא בדמיון בצורות צימוח, תכונות מורפולוגיות, מאפייני ה- life history או בתהליכים-מנגנוניים פיזיולוגיים. הקונברגנציה עוסקת, אם כך, במבנה ובתפקוד של האורגניזם ולא במצבו הגנטי-סיסטמתי.
המיון הקלאסי של חברות ויחידות צומח בראשית המאה התבסס על הנחת הקונברגנציה: תנאי אקלים מסוימים יביאו להתפתחות חברה מסוימת, ובכל מקום שישררו תנאים דומים תתפתח חברת צומח דומה. ניתן לראות במודל של Holdridge (1947 ) את תמצית התיאוריה. תצפיותיו וקביעותיו של Schimper (1903) על הצומח הים תיכוני תאמו את התפיסה הרווחת. הוא טען שבמוקדי האקלים הים-תיכוני השונים מתפתחות חברות בעלות מבנה ותכונות (סקלרופיליות, שורשים מפותחים) דומות, אפילו שאין כמעט קירבה בהרכב הפלוריסטי של החברות. מאז ימי שימפר, נעשו ניסיונות רבים להשוואה בין יחידות צומח ים תיכוני שהתפתחו במנותק באזורים שונים (Shmida, 1981; Naveh, 1967 ו- Shmida and Barbour,1982 השוו בין ישראל וקליפורניה; Specht, 1969 השווה בין אוסטרליה, צרפת וקליפורניה ו- Mooney et al., 1970 ו-Mooney and Dunn, 1970 השוו בין צ'ילי וקליפורניה). השוואות אלה התמקדו במאפיינים מבניים ופיזיולוגיים והראו קווי דמיון בולטים בין החברות השונות.
בשנות ה-60 וה-70, עלה נושא הקונברגנציה של אקוסיסטמות (חברות האורגניזמים + הסביבה הפיזיקלית). ניסיון זה נתקל בקשיים תיאורטיים ולא פותח.
ניתוח בוטני, התומך בתיאוריית הקונברגנציה לגבי צומח ים-תיכוני בצ'ילי וקליפורניה, ניתן על ידי פארסונס וחובריו (Parsons, 1967; Parsons and Moldenke, 1975). הוא מראה שצורות החיים והמורפולוגיה של העלים במיני
צמחים שאינם קרובים, לאורך גרדיאנטים סביבתיים בשתי היבשות, דומים מאוד בבתי גידול דומים. הדמיון הזה גבוה מהדמיון שבין צמחים מאותן משפחות, או מאותו סוג, הגדלים בבתי גידול שונים באותה יבשת. אבל, כאשר הורחבה מסגרת ההשוואה, והכילה גם את צמחיית האזור הים-תיכוני בכף הדרום אפריקאי, לא נמצאו עדויות תומכות לקונברגנציה. צמחיית ה- fynbos הדרום אפריקאי שונה בעושר הפלוריסטי, צורת הצימוח, גודל ואורך חיי העלים ובקוצניות המינים המעוצים מצמחיית צ'ילי וקליפורניה (1980 ,Campbell and Cowling). ההסבר שניתן להבדלים אלה היה הקרקעות העניות עליהן התפתח הצומח הים-תיכוני בדרום אפריקה. Axelrod הראה שינויים דומים בתכונות המורפולוגיות של עלים סקלרופיליים לאורך גרדיאנט סביבתי בקליפורניה ובאגן הים התיכון. כללית ניתן לומר, שטיפוס העלים הוא התכונה המאפיינת את שיחי כל האזורים הים-תיכוניים: רובם ירוקי-עד עם יחס גבוה של משקל לשטח (סקלורפיליות). גם מקצב הפוטוסינתיזה דומה בין הצמחים ביבשות השונות (Cody and Mooney, 1978). הסקלרופיליות מוסברת כהתאמה לתנאי מחסור עונתי במים ו/או התאמה לקרקעות עניות, בעיקר בזרחן (1967 ,Loveless).
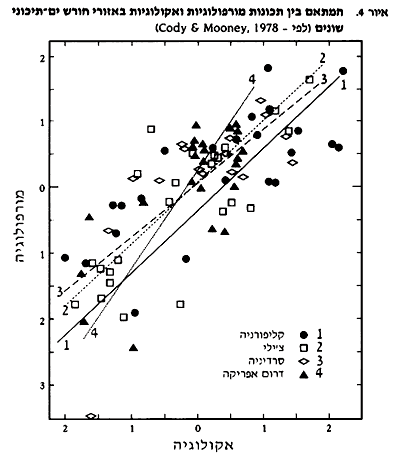
דמיון מורפולוגי ואקולוגי נמצא גם בין בעלי חיים המאכלסים את החורש הים-תיכוני ביבשות השונות. cody ו- Mooney (1978) הראו, בעזרת ניתוחים סטטיסטיים, כי לשונות המורפולוגית של ציפורי חורש היו ערכים ממוצעים, טווח ופיזור דומים ביבשות השונות. גם טווח הגבהים בהם אכלו הציפורים, והתנהגות האכילה היו דומים. זאת, כאשר אין דמיון סיסטמתי בין מרבית מיני הציפורים. ניתוח המתאם בין תכונות מורפולוגיות לאקולוגיות הראה גם הוא דגם דומה בין היבשות (ראה איור 4), כאשר חוזק המתאם עשוי להשתנות לפי התכונה האקולוגית הנבדקת. גם בתכונות חברת בעלי החיים, כמו: מספר המינים, פיזור המינים בין הרמות הטרופיות וצפיפות המינים בעלי תיפקוד אקולוגי דומה, נמצא דמיון.
איור 5 מראה על הקשר שבין גובה הצומח וצפיפותו ובין גובה הצומח ופיזור הציפורים והחרקים בו, ב-4 מוקדי התפתחות של חורש ים-תיכוני בעולם.

בצד קווי דמיון סביבתיים (אקלים) ומורפולוגיים-אקולוגיים (צמחים ובע"ח) יש גם לא מעט נקודות שוני בין תצורות החורש השונות בעולם. גם איפיון מדויק יותר של האקלים מראה הבדלים, בעיקר באופי הקיץ (ראה איור 1). ההרכב הפלוריסטי שונה כמו גם מבנה חברות הצמחים, היחסים הסוקצסיוניים ומקצב הצמיחה של הצמחים השליטים. שוני בזמינות המשאבים יוצר הבדלים במספר מיני בעלי החיים וצפיפותם ומביא לשונות גבוהה מהצפוי של הרכב החברה.
טבלה 3 מסכמת את הדמיון והשוני בין האזורים הים-תיכוניים (Cody and Mooney, 1978).

התפתחות קונברגנטית מוגבלת על ידי שונות בתנאים האקולוגיים בכל אתר, המבנה הגנטי הבסיסי של האורגניזמים המתפתחים, טווח זמן מספיק שיאפשר התפתחות כזו ואירועים היסטוריים מקריים או מעשה ידי אדם, שיכולים היו להשפיע על מהלך ההתפתחות הטבעית (Cody and Mooney, 1978).
בשנים האחרונות, ככל שנאספים נתונים מדעיים מסודרים על הצומח באזורים הים-תיכוניים השונים, עולה ביקורת מהותית על רעיון הקונברגנציה הים-תיכונית. בארבור ומיניק סיכמו לאחרונה נתונים אלה וגיבשו מניפסט ביקורתי תחת הכותרת "המיתוס של הקונברגנציה בחורש הים-תיכוני" (Barbour and Minnich, 1990). הם טוענים שלמרות שקיים דמיון מורפולוגי גבוה בין תצורות הצומח באזורים הים-תיכוניים השונים, הוא למעשה חלקי מאוד ושטחי, בעוד שההבדלים רבים מאוד ומשמעותיים. המחברים מציגים רשימה ארוכה של הבדלים:
- הקרקעות שונות במבנה הכימי. באוסטרליה ודרום-אפריקה (ד"א) הן עניות ביותר: ריכוז חנקן וזרחן כ- 0.0001%, בעוד שבאזור אגן הים התיכון (אי"ת) חנקן מהווה 0.4% וזרחן - 0.03%.
- העוצמה של היובש הקיצי שונה. בצ'ילי לא יותר מ- 5% מהמשקעים השנתיים יורדים ב- 4 חודשי קיץ בעוד שבאוסטרליה וד"א יורדים 15% בזמן זה.
- המשרעת השנתית של הטמפרטורה שונה מאוד. ההפרש בין הטמפרטורה היומית הממוצעת בחודש החם ביותר לטמפרטורה היומית הממוצעת בחודש הקר ביותר - בקליפורניה ואי"ת הוא כ- 27°C ובאזורים האחרים כ- 17°C.
- נוכחות זרם מים קר ליד החופים. באוסטרליה ואי"ת אין זרם כזה ובשאר האזורים הוא קיים ונוכחותו מביאה ערפל עונתי בעל השפעה אקלימית ממתנת.
- לחות אוויר בקיץ. באגן הים התיכון אקלים לח בקיץ, בשונה משאר האזורים.
- עושר מיני הצומח. בד"א יש פי 5-3 מינים מאשר בקליפורניה, ואילו לשאר האזורים יש ערכי ביניים.
- הרכב צורות החיים. באוסטרליה צומח מעוצה מהווה 63%-58% מהפלורה, ובצ'ילי רק 21%.
- היחס בין 2 הקבוצות העשבוניות: חד- ורב-שנתיים משתנה. בד"א ואוסטרליה רק 5%-2% מהפלורה הם חד-שנתיים בעוד שבאי"ת ובצ'ילי - מעל 30%.
- המורפולוגיה של העלים שונה. בד"א יש נוכחות גבוהה של בעלי עלים קטנים (קטן מ-25 מ"מ) בצ'ילי ואי"ת יש מעט מינים כאלה.
- נוכחות נשירי חורף או נשירי-יובש: גבוהה באי"ת ובצ'ילי לעומת קליפורניה.
- קוצניות (עלים או גבעולים): נפוצה באי"ת וחסרה לחלוטין בד"א.
- פיזור הביומסה העל-קרקעית: בד"א גובה מקסימלי - 3.5 מ', וחצי מהביומסה מופיע בגובה נמוך מ-1.5 מ', בקליפורניה הגובה עד 7 מ', וחצי מהביומסה גבוהה מ- 2 מ'.
- כיסוי צומח מעוצה. באוסטרליה, קליפורניה וד"א הכיסוי רציף. באי"ת וצ'ילי יש בו מפתחים וקרחות ובהם צומח עשבוני (לכן בקליפורניה רק 1% של כיסוי עשבוני לעומת 20% ומעלה בצ'ילי.)
- ביומסה על-קרקעית: נעה בין 750 גר'/מ"ר בצ'ילי ל-2,300 גר'/מ"ר בקליפורניה ואי"ת.
- ייצור ראשוני נטו: מ-250 גר'/מ"ר/שנה בצ'ילי ועד 670 גר'/מ"ר/שנה בקליפורניה ואי"ת.
- שיא עונת הצימוח העשבוני: אביב בחצי הדרומי, בעוד שבחצי הצפוני יש 2 תקופות צימוח (אביב וקיץ).
- זמן ותיארוך פעילות דרסטית של האדם: באי"ת וצ'ילי, האדם הלבן פעל באינטנסיביות, אם כי באגן הים התיכון החלה הפעילות זמן רב (אלפי שנה) קודם. באוסטרליה וקליפורניה פעילות כמעט ולא הורגשה והחלה רק בתקופה מאוחרת.
- תדירות שריפות: תדירות גבוהה בד"א (מחזור 30-6 שנה) בקליפורניה ובצ'ילי.
- תגובת הצומח לשריפה: לא קבועה וברורה!
- ליגנוטוברס (Lignotubers) (חלקים מעוצים של הגבעול התחתון, ולפעמים של החלק העליון של השורש, המשמשים אברי התחדשות ואגירה, בעיקר במינים ופרטים הנמצאים תחת משטר של מחזורי שריפה) נפוץ יותר באוסטרליה וקליפורניה.
המסקנה: ההבדלים רבים מאשר הדמיון. חלק מההבדלים יכול לנבוע מאורך ואינטנסיביות פעולת האדם; או בגלל המבנה (ה"מחויבות") הגנטי הראשוני.
א. האקלים הים-תיכוני
ג. החורש באגן הים התיכון
ד. החורש הים-תיכוני בישראל
ה1. התאמות של צמחי החורש לסביבה (חלק 1)
ה2. התאמות של צמחי החורש לסביבה (חלק 2)
ה3. התאמות של צמחי החורש לסביבה (חלק 3)
ו1. השפעת האדם על הצומח הים-תיכוני (חלק 1)
ו2. השפעת האדם על הצומח הים-תיכוני (חלק 2)
ז. ממשק החורש הים-תיכוני
| ביבליוגרפיה: | ||||||||||||||
|
|
|
חזרה3 |
 |
הדפסה | ||||||
| החומר במאגר זה הינו לשימוש פרטי ולשימושם של מורים ותלמידים לצרכים לימודיים בלבד. אין להפיץ, להעתיק, לשדר או לפרסם חומר כלשהו מתוך המאגר, ללא הסכמה מראש ובכתב של בעלי זכויות היוצרים השונים, המצויינים בתחתית כל פריט. | |||||||
|
|||||||
| החומר במאגר זה הינו לשימוש פרטי ולשימושם של מורים ותלמידים לצרכים לימודיים בלבד. אין להפיץ, להעתיק, לשדר או לפרסם חומר כלשהו מתוך המאגר, ללא הסכמה מראש ובכתב של בעלי זכויות היוצרים השונים, המצויינים בתחתית כל פריט. |